Saif Rasool1, Afaq Ahmad 2 and Mohammad Ashraf2*
1 School of Biological Sciences, University of the Punjab, Lahore, Pakistan
2 Gulab Devi Chest Hospital, Lahore, Pakistan
3 Nuffield Department of Medicine, University of Oxford, OX37BN, United Kingdom
4 Department of Pathology and Laboratory Medicine, University of California, Davis, California, United States of America
5 School of Biochemistry and Biotechnology, University of the Punjab, Lahore, Pakistan
PJEST. 2023, 4(3); https://doi.org/10.58619/pjest.v4i3.109 (registering DOI)
Received: 12-April-2023 / Revised and Accepted: 23-June-2023 / Published On-Line: 28-June-2023
ABSTRACT: Tuberculosis (TB) an opportunistic disease still remains a major cause of death globally after COVID-19 pandemic and HIV. Enhanced microbial resistance to therapeutic drugs has paved way to introducing new vaccines for prophylactic control of the disease. Timely diagnosis of TB is also highly needed for effective treatment. Therefore, a fast reliable diagnostic method with high sensitivity is need of the day. In our study, CFP10 and ESAT-6 antigens were cloned, expressed, and purified individually and as a chimeric construct in Escherichia coli BL21. Epitopic analysis revealed that the CFP10-ESAT-6 (CEP) chimeric construct may be more sensitive than the individual sensitivities of CFP10 and ESAT-6. Moreover, molecular analysis of secondary structure and 3-D modelling of each construct also validated that CEP may prove a better immunodiagnostic tool in addition to previously reported EC skin test.
Key Words: Tuberculosis, Novel epitope prediction, EC Skin test, CEP Fusion, Diagnostic potential
Introduction
Tuberculosis (TB) continues to be the second most serious infectious illness even during corona epidemic. The WHO End TB Strategy’s main objectives could not be met due to a significant reduction in TB surveillance which increased the mortality toll. A decline in treatment and greater number of fatalities can be attributed to the approximately 20.2 million people of all ages who are still untreated worldwide [1]. Pakistan is still one of the eight nations that will make up two-thirds of the thirty nations with the highest TB burden in 2020. A more accurate, reasonably priced, and optimally scaled-up diagnostic test is crucial for the disease’s effective control. An overly sensitive serodiagnostic test can aid in early detection in countries with a high TB incidence, such as Pakistan.
In silico evaluation of both new and old vaccines is aiding in the formulation of innovative vaccine designs [2]. Recently, a number of computational techniques have been created that evaluate the immunogenic components of microbes using genomic sequence data. These regions, often referred to as epitopic regions, are the best sections of vaccinations [3]. The expression of Mtb genes has been improved when the whole Mtb genome sequence was grasped using extremely specific and sensitive methods [4].
Understanding how environmental changes affect the regulatory gene activities of Mtb will aid in understanding the early, latent, and resuscitation stages of TB [5]. Combining in-vivo and in silico research enables the creation of vaccines to concentrate on a smaller set of pertinent antigens [6]. We chose Mtb genes that co-express in vivo in a pool of antigens.
Many of the highly expressed Mtb genes relate to the DosR regulon [7] which is in charge of the transcriptional alterations brought on by oxygen deprivation, which is thought to be a significant trigger for Mtb to enter a dormant state [8]. EHR, which is separate from DosR, comprises of a bigger set of genes that are strongly activated in response to hypoxia for longer periods of time [9]. Additionally, EHR exhibits a significant overlap with Mtb genes that are brought on by food deficiency [10]. The equipment needed to enter and endure delay may be present in the EHR [11].
The reactivation of intracellular tubercle bacilli while the host’s immune system is being suppressed is a crucial stage of tuberculosis. Mtb has five genes that significantly resemble Micrococcus luteus’ resuscitation promoting factor (Rpf). Rpfs are tiny proteins that are present in a variety of gram-positive, high G/C organisms. The Rpf proteins of Mtb have been demonstrated to boost the growth of extended-stationary-phase cultures of BCG in vitro at picomolar concentrations. Rpfs may be involved in the in-vivo resuscitation action. Human tissues infected with Mtb have recently been found to express these proteins [12] & [13].
Epitope-based diagnostics and vaccinations clearly require the discovery of epitopes suitable for therapeutic or preventive action as well as for diagnostic application. Numerous anticipated epitopes have since undergone experimental confirmation, proving the value of such predictions. Several distinct algorithms, including PREDITOP, PEOPLE, BEPITOPE, and BcePred, have been developed for predicting B-cell epitopes based on the physico-chemical characteristics of amino acids. The B-EpiPred server was used in our investigation to identify promiscuous B-cell epitopes of two highly expressed Mtb proteins in vivo [14].
The service locates opportunistic B-cell linear epitopes that can trigger an appropriate immunological response in several populations throughout the world. The fact that these techniques are qualitative rather than quantitative is their main flaw. Finding the precise start and end residues that potentially behave as B-cell epitopes is extremely difficult [15]. For the prediction of B-cell epitopes of Mtb chosen proteins, we integrated two versions of the server, B-EpiPred-1 and 2. The two techniques were combined to make it easier to choose extremely precise B-cell epitopes in each protein sequence. For the creation of a vaccine, choosing areas with both T- and B-cell epitopes may be crucial. Both humoral and cell-mediated immune responses, which are both crucial for preventing Mtb, are anticipated to be elicited by these epitopes. It is well known that Mtb is an intracellular pathogen, and that the immune system’s ability to fight off bacterial infections is greatly influenced by cell-mediated immunity. As a result, a lot of research on creating new T-cell epitope-based vaccinations against TB also focuses on this [16].
Single native antigens have not been able to reliably produce consistent sensitivity and specificity in TB serodiagnosis. Practical and affordable TB serodiagnosis can be achieved by the development of more sensitive fusion antigens with improved epitope availability. In our labs, various special TB antigen combinations (full length and shortened) were created for efficient antibody-based diagnostics, and their diagnostic utility was assessed against a broad spectrum of people.
“Virulence-associated type VII secretion system” is a part of (RD1). The current BCG strains in use, lack this region and hence the secretion system. This deletion results in loss otherwise a strong immunological reaction [17]. The two significant secretory antigenic proteins Rv3874 (CFP10) and Rv3875 (ESAT-6) are part of this system. They exhibit pathogenicity in pathogenic mycobacteria, which is well supported by the available evidence [18]. Therefore these two antigens are candidates for vaccine development as well as in serodiagnosis. MTBVAC which consists of live attenuated Mycobacterium tuberculosis includes the RD1 region [19].
DIVA test also includes a recombinant fusion Rv3875-Rv3874 in a cocktail of peptides. This test is also used for animals with and without TB infection after vaccination.
In contrast, we have designed fusion construct CEP (named as CEP after the names of Culture filtrate protein 10 and early secretory antigenic target-6). Our CEP fusion is currently under study in our lab for immunological characterization using sera from TB-affected and healthy people in the Pakistani community.
Materials and Methodology
Epitope prediction
The full-length native CFP10 and ESAT-6 antigens were used in this study. The reported epitopes were taken from IEDB [20]. The Bepipred-1.0 Linear Epitope Prediction tool (http://tools.iedb.org/bcell) was used for possible B cell epitopes of ESAT-6 [21] . As seen in the graph of residue placements versus residue outcomes in Figure 2a, a threshold of 0.35 was chosen. Seven total epitopes, including those extracted from IEDB, were chosen for the fusion with a threshold length of ten. The positions of the amino acids and the length of the sequence agreed with the outcomes of the Bepipred 1.0 linear epitope prediction. Software called Bepipered-2 was also utilised to evaluate any growth in the quantity or range of active epitopes in the fusion (Fig. 2 b–d).
Cloning, Expression, and Purification
Using the genomic DNA of the Mtb strain H37Rv, the CFP10 (303 bp) and ESAT-6 (288 bp) genes were amplified [22]. For CFP10, pF1 and pR1 primers were utilized, whereas for ESAT-6, pF2 and pR2 primers were employed (Table 1). The PCR programme conditions used to amplify CFP10 and ESAT-6 are listed in Table 2. After cloning into pJET1.2, E. coli DH10 cells were subjected to transformation. As previously described [23], sub-cloning was done using pET28a (+), and then the CFP10 and the chimeric pET28a (+) were double digested with NdeI and BamHI, whereas ESAT-6 was digested with BamHI and HindIII. Products of digestion were ligated and transformed into DH10 cells.
NdeI and BamHI digestion of amplified CFP10 without a stop codon and pET28a (+)-ESAT-6 was followed by mutual ligation to create pET28a (+)-CFP10-ESAT-6 (CEP). The double digestion analysis and sequencing confirmed the success of cloning the antigenic genes.
Table 1 Primer details for genes selected from MTB Genome (colored nucleotides represent the restriction enzyme recognition sites)
| Selected gene with size | Title | Optimized primer Sequence 5‘-3‘ | Restriction Enzymes |
| Rv3874 (303bp) | pF-1 | AGAGCATATGGCAGAGATGAAGACCGA | NdeI |
| pR-1 | ATTAGGATCCGAAGCCCATTTGCGAGG | BamHI | |
| Rv3875 (288bp) | PF-2 | ATATGGATCCATGACAGAGCAGCAGTG | BamHI |
| PR-2 | ATATAAGCTTCTATGCGAACATCCCAGTGA | HindIII |
Table 2 PCR Profile for individual genes
| Antigens | Initial Denaturation | Number of cyclic repeats= 30 | Final Extension | ||
| Denaturation | Annealing | Extension | |||
| Rv3874 & Rv3875 | 95°C (5 minutes) | 95°C (45sec) | 66°C (45sec) | 72°C
(1 minute) |
72°C (8 minutes) |
The CEP fusion protein (25kDa) was expressed at the IPTG induction concentration of 0.1 mM in BL21 cells. The cells were then spun and re-suspended in Tris buffer. Sonication was then used to disrupt the cells. After centrifugation for 12 minutes at 4500 rpm, the components were resolved on SDS-PAGE. According to previously published techniques, all the histidine-tagged recombinant proteins were purified. The concentration and purity were determined by Bradford assay and SDS-PAGE analysis respectively. Dialysis with 20 mM Tris-Cl (pH 8.0) was used to remove the salts from the eluted fractions.
In silico structural analysis
Protein molecular modelling was carried out using the Raptor-X Server [24]. Protein secondary and tertiary structures are predicted, together with solvent accessibility, a distance map with enhanced alignment accuracy. It is a modelling tool with templates. As previously indicated, chimeric models were created using a variety of bioinformatics tools and servers, followed by calculation of validation scores, hydropathy and model visualisation [25]. Additionally, the Ramachandran plot statistical analysis was carried out to identify advantageous residues for fusion.
Evaluation of therapeutic ability
All of the recombinant antigens’ vaccine potential was evaluated favourably using the online prediction engine Vaxijen.
Results
Epitopes of native and fusion proteins
A qualitative binding assay was used to identify one epitope for the antigenic protein Rv3874 the sequences shown in table 3.1 were obtained in silico. The number of predicted epitopes for Rv3875 were two (See table 3.1). There are no known epitopes for this protein have been identified so far.
Table 3.1 Predicted epitopes of individual antigens
| ESAT-6 epitopes with position | CFP10 epitopes with position (two of four shown here) |
| AWGGSGSEAYQGVQQKWDATATE ((42–64) | EMKTDAATLAQEAGNFER (3-20) |
| SEAGQAMASTEGNV (77–90) | DQVESTAGSLQGQWRGAAGTAA (30-51) |
Complete sequence of CFP10 and ESAT-6 genes were joined to create a construct that contained all of the aforementioned epitopes (Figs. 2 and 3). Using the programme Bepipred-2, the locations of the epitopes in the two distinct antigens and their fusion were also examined (Figure 2b–d). These predictions are based on protein linear sequence. Therefore this software showed that fusion of the two antigens enhanced the epitope at the N terminal residues of CFP10 (5-20). Additionally, the fusion molecule’s N terminal residues of Rv3875 (1–7) and CFP10 (97–100) became a member of the active epitopic area (Figure 2d).
Cloning, expression, and purification
SDS-PAGE was used to analyse the proteins, and the protein bands that were thus resolved had the predicted sizes. 90 percent of CEP was expressed. While the rest produced soluble expression, Rv3875 was only moderately soluble. A nickel column purification process produced a 98% pure fusion protein (Fig. 1). After purification, 47% of CEP was recovered.

Fig. 1. recombinant fusion expressed and purified shown on SDS PAGE gel: 1– un-induced culture, 2– induced culture at 0.1mM IPTG, 3 shows Supernatant 4 shows pellet, 5 Protein marker and 6 shows purified fusion.
Computational analysis of structure
Recombinant protein 3D molecular models were created using the Raptor-X server. Using the previously reported tools and software, the computational protein models showed high hydropathy scores [26], (Fig. 4 (a-f). The same was also supported by Ramachandran plot data. It revealed 170 residues (96% of the amino acids) in the most advantageous region, 3.4% in extra acceptable regions, and 0% in banned regions. In terms of maximum amino acid exposure, the plot exhibits a 2 Angstrom resolution and R factor >20, making it a potent antigen (Fig. 5).

Fig. 2. (a) Comparison of prediction results for B-cell epitopes of fusion antigen (b) comparison of CFP10, ESAT-6 epitopes and epitopes of their CEP fusion.

Fig. 3. Construction map of Fusion CEP from native antigens with number and position of antigens for schematic details. Two extra amino acids were added represented by GS or black bar at the point of fusion. These amino acids were added as a result of BamHI restriction site insertion and CEP fusion is showing cumulative effect of epitopes in the fusion.

Fig. 4. (a-f) Ribbon models in 3D with epitopes showing solvent accessibility of cfp10, ESAT-6 and their fusion CEP. Epitopes in left figure are shown in black color whereas responsive amino acids in the right figure are labelled with red color depicts along with green color which shows supporting effect for antigen-immunoglobin interaction. The non-supporting residues are shown in blue color.
Assessment of vaccine potential
Vaxijen, an online prediction tool used for in silico assessment of prospective prophylactic efficacy of antigens in the creation of vaccine, estimated the vaccination potentials of the native as well as the fusion constructions. CEP scored at 0.67 with a threshold of 0.4, indicating a high potential for a vaccine. This positive finding strengthened the theory that amino acids should be arranged in a better way to mount an improved immune response.
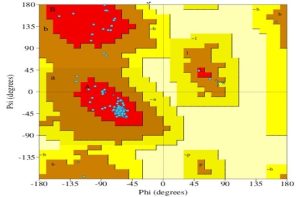
Fig 5: Ramachandran statistical analysis plot for CEP fusion where the part of graph shown in red color represents amino acid residues in a position most suitable for inter action. The additionally allowed regions of amino acid residues are shown in yellow color and the residues which have slight effect on immunogenesis are shown in light yellow color. The residues with no role are shown white.
Discussion
In countries like Pakistan with limited economic resources and high TB incidence, TB serodiagnosis using chimeric fusion combinations can be cost-effective and practical. To do this, multistage antigens may incur higher expenditures. Therefore, it will be less expensive to create fusion constructions from various antigens to improve the diagnostic effectiveness of Enzyme Linked Immunosorbent Assay. However, a size range that is unstable may occur from the merging of several full-length genes. Therefore, in this study, the fusion molecule was created using paired antigens. Rv3874-Rv3875 chimeric protein was observed to be overexpressed when Rv3875’s C terminus was fused to Rv3874’s. The improved concentration of fusion to 300mg/L, was also a result of fusing cfp10 at the N-terminal.
Regardless of the specific infection stage, the symptoms in the patient’s history point to the invasive infection stage. Before any pharmacological intervention began, serum samples were taken. It indicates that in certain conditions, the patient’s sera will produce enough antibodies as a result of a continuous immunological response.
Epitopic residues that are accessible to solvents will interact and be exposed to antibodies more effectively [27]. The Consensus prediction interface residues in transient complexes service employs molecular simulations for solvent accessibility studies, confirming the idea that the fusion of two antigenic proteins can enhance the alignment of the epitopes through additive mechanisms. This will improve the antigen-antibody interaction’s efficiency and increase its sensitivity. The CEP fusion would have a combined sensitivity greater than the sum of individual sensitivities of the two constituent antigens keeping in view the neo-epitopic peptide generation. The antigens’ structural reorientation when in the fusion state is the cause. The development of a more potent antigenic epitope at Rv3874’s N terminal (5-20) and its C terminal residues (97–100), which were epitopically more active in conjunction with N terminal residues (1–7) of Rv3875 (Fig. 1), further corroborate this theory.
Our fusion Rv3874-Rv3875 may show better solubility which facilitates chromatographic purification. Therefore, it is expected that CEP may provide a significant chance of developing into a quick, reliable, and affordable serodiagnostic tool for TB
Conclusions
The current study represents a novel in silico approach for the identification of antigenic regions. The selected candidates may prove to be both vaccine and diagnostic candidates for TB. Creating chimeric antigenic proteins is more practical and affordable than synthesizing bulk peptides. The CEP fusion construct could be a turning point in the creation of other chimeric molecules with improved serodiagnostic capabilities. The combination of antigens and the potential development of promiscuous peptides is expected to increase the sensitivity of the chimeric CEP protein. In contrast, the RV3875’s attachment produced a more advantageous orientation with increased sensitivity. However, these are just preliminary results, and further research is still required to confirm actual CEP fusion applications.
Ethical approval
Ethical guidelines regarding patient care, as well as the use of experimental animals, were approved by the Ethics Committee, School of Biological Sciences, University of the Punjab, Lahore, Pakistan vide letter no. SBS/765/17 and followed accordingly.
Funding source
Pakistan Academy of Sciences Grant No. 59/PAS/1269 and the research support fund of Indigenous Ph.D. Scholarship (Higher Education Commission of Pakistan) was used to support this research work.
Author contribution
Conceptualization, resource contribution, and overall supervision were done by M.WA., N.M., performed major experiments and data analysis. Demographical parameters and plasma samples of volunteers were recorded by Z.U.N., and I.H.K., Critical reading and draft writing were done by N.M., N.H., U.R., H.K., and M.W.A.,
Declaration of competing interest
None.
Acknowledgments
We are grateful to the medical staff at Gulab Devi Chest Hospital, Lahore for the collection of plasma samples from TB patients and healthy subjects.
References
[1] M. Reid, S. Silva, and E. P. Goosby, “World TB day: a new opportunity to reimagine workplace health,” BMJ Global Health, vol. 6, no. 3, p. e005601, 2021.
[2] F. Zhou, X. Xu, S. Wu, X. Cui, L. Fan, and W. Pan, “Protein array identification of protein markers for serodiagnosis of Mycobacterium tuberculosis infection,” Scientific reports, vol. 5, no. 1, p. 15349, 2015.
[3] Z. Yan et al., “Evaluation of panels of Mycobacterium tuberculosis antigens for serodiagnosis of tuberculosis,” The International Journal of Tuberculosis and Lung Disease, vol. 22, no. 8, pp. 959-965, 2018.
[4] M. Afzal, S. Khurshid, R. Khalid, R. Z. Paracha, I. H. Khan, and M. W. Akhtar, “Fusion of selected regions of mycobacterial antigens for enhancing sensitivity in serodiagnosis of tuberculosis,” Journal of microbiological methods, vol. 115, pp. 104-111, 2015.
[5] M. Akhter, S. Arif, A. Khaliq, Z. un Nisa, I. H. Khan, and M. W. Akhtar, “Designing fusion molecules from antigens of Mycobacterium tuberculosis for detection of multiple antibodies in plasma of TB patients,” Tuberculosis, vol. 124, p. 101981, 2020.
[6] S. Sulman et al., “Enhanced serodiagnostic potential of a fusion molecule consisting of Rv1793, Rv2628 and a truncated Rv2608 of Mycobacterium tuberculosis,” Plos one, vol. 16, no. 11, p. e0258389, 2021.
[7] S. Arif, M. Akhter, A. Khaliq, Z. un Nisa, I. H. Khan, and M. W. Akhtar, “Serodiagnostic evaluation of fusion proteins from multiple antigens of Mycobacterium tuberculosis for active TB,” Tuberculosis, vol. 127, p. 102053, 2021.
[8] S. Khurshid, M. Afzal, R. Khalid, I. H. Khan, and M. W. Akhtar, “Improving sensitivity for serodiagnosis of tuberculosis using TB16. 3-echA1 fusion protein,” Tuberculosis, vol. 94, no. 5, pp. 519-524, 2014.
[9] S. Khurshid, R. Khalid, M. Afzal, and M. W. Akhtar, “Truncation of PstS1 antigen of Mycobacterium tuberculosis improves diagnostic efficiency,” Tuberculosis, vol. 93, no. 6, pp. 654-659, 2013.
[10] A. M. Abdallah et al., “Type VII secretion—mycobacteria show the way,” Nature reviews microbiology, vol. 5, no. 11, pp. 883-891, 2007.
[11] M. Behr et al., “Comparative genomics of BCG vaccines by whole-genome DNA microarray,” Science, vol. 284, no. 5419, pp. 1520-1523, 1999.
[12] L. Chen et al., “The development and preliminary evaluation of a new Mycobacterium tuberculosis vaccine comprising Ag85b, HspX and CFP-10: ESAT-6 fusion protein with CpG DNA and aluminum hydroxide adjuvants,” FEMS Immunology & Medical Microbiology, vol. 59, no. 1, pp. 42-52, 2010.
[13] M. E. Sarmiento et al., “Immunoinformatics study on highly expressed Mycobacterium tuberculosis genes during infection,” Tuberculosis, vol. 94, no. 5, pp. 475-481, 2014.
[14] G. J. Jones, A. Whelan, D. Clifford, M. Coad, and H. M. Vordermeier, “Improved skin test for differential diagnosis of bovine tuberculosis by the addition of Rv3020c-derived peptides,” Clinical and Vaccine Immunology, vol. 19, no. 4, pp. 620-622, 2012.
[15] H. Köhler et al., “Immune reactions in cattle after immunization with a Mycobacterium paratuberculosis vaccine and implications for the diagnosis of M. paratuberculosis and M. bovis infections,” Journal of Veterinary Medicine, Series B, vol. 48, no. 3, pp. 185-195, 2001.
[16] S. H. Kaufmann et al., “Progress in tuberculosis vaccine development and host-directed therapies—a state of the art review,” The lancet Respiratory medicine, vol. 2, no. 4, pp. 301-320, 2014.
[17] T. Hsu et al., “The primary mechanism of attenuation of bacillus Calmette–Guerin is a loss of secreted lytic function required for invasion of lung interstitial tissue,” Proceedings of the National Academy of Sciences, vol. 100, no. 21, pp. 12420-12425, 2003.
[18] N. Aguilo, S. Uranga, D. Marinova, M. Monzon, J. Badiola, and C. Martin, “MTBVAC vaccine is safe, immunogenic and confers protective efficacy against Mycobacterium tuberculosis in newborn mice,” Tuberculosis, vol. 96, pp. 71-74, 2016.
[19] N. Aguilo et al., “Reactogenicity to major tuberculosis antigens absent in BCG is linked to improved protection against Mycobacterium tuberculosis,” Nature communications, vol. 8, no. 1, p. 16085, 2017.
[20] R. Vita et al., “The immune epitope database 2.0,” Nucleic acids research, vol. 38, no. suppl_1, pp. D854-D862, 2010.
[21] R.-P. T. Larsen and A. Studentlitteratur, “Lund,” ed: Sweden, 2003.
[22] R. Khalid, M. Afzal, S. Khurshid, R. Z. Paracha, I. H. Khan, and M. W. Akhtar, “Fusion molecules of heat shock protein HSPX with other antigens of Mycobacterium tuberculosis show high potential in serodiagnosis of tuberculosis,” PloS one, vol. 11, no. 9, p. e0163349, 2016.
[23] J. Sambrook, “Plasmid and their usefulness in molecular cloning,” Molecular cloning, a laboratory manual, vol. 1, pp. 1.31-1.162, 2001.
[24] M. Källberg et al., “Template-based protein structure modeling using the RaptorX web server,” Nature protocols, vol. 7, no. 8, pp. 1511-1522, 2012.
[25] P. C. Hopewell, M. Pai, D. Maher, M. Uplekar, and M. C. Raviglione, “International standards for tuberculosis care,” The Lancet infectious diseases, vol. 6, no. 11, pp. 710-725, 2006.
[26] G. Scarabelli, G. Morra, and G. Colombo, “Predicting interaction sites from the energetics of isolated proteins: a new approach to epitope mapping,” Biophysical journal, vol. 98, no. 9, pp. 1966-1975, 2010.
[27] S. J. de Vries and A. M. Bonvin, “CPORT: a consensus interface predictor and its performance in prediction-driven docking with HADDOCK,” PloS one, vol. 6, no. 3, p. e17695, 2011.